I would like to thank Ivan Gomez.

---
Teleology as applied to the study of biological evolution has a history going back to Plato and Aristotle. Plato attempted to ground final causes into those necessary ends which were the good based on the whim of an external agency (9; sec. 99), whereas Aristotle grounded his account as a type of formative force intrinsic to natural forms (10; pp. 639-643). He [Aristotle] also presents four causes in his Physics. One of them, the efficient cause, is the principle which, as I will show, begins with the primitive entropic relation between the original components which have been implicated in the origin of life. The final ends assigned to biological structures constituting organisms have become part of a linguistic game given by the statement S: “the structure X is for the function Y” (5; line 1). I use the term biological structure to mean those components which we can view under the microscope that seem to constitute organic life when cultured – morphological structures. Indeed, this view has served well in the fields of Medicine and Microbiology (e.g. “Genes are for the encoding of proteins”). However, it has failed to provide an account for the phenomena underlying the unity of living organisms due to its inability to achieve a complete model encompassing the plurality of functions which biological structures are capable of expressing within the constraints of this Universe’s physics, the world which we can observe around us. Consider the case of a mousetrap. We would say that its function – the natural purpose that comes from its structure – is to catch mice. What is not considered is that nothing would stop me from using it as a tie-clip, either. This example elucidates what is meant by the plurality of functions that the organism is capable of expressing, whether it is the components making it up or the whole organism interacting with the environment.
Mechanistic theories that explain highly organized systems as having a list of structures working towards natural purposes cannot be explained without reference to immanent teleology (6; pp. 22). When a mechanism or otherwise linear pathway is proposed for the function of an organism, the classical teleology would ascribe this generally as S. S can further be extended in a chain of causation going backwards and forward in time and space. The structure X, initiating function Y which enacts on structure Y’ and so on, is the backbone of such mechanisms or bauplan/wiring diagrams for organisms, tracing the chain from the birth of the living system to its destruction. From the DNA/RNA come proteins, functional units which are used for building blocks involved in a hierarchy of structures going from the single cell up to entire organs, which compose the organism. Mind you, this only applies for the case of organisms which transfer a genetic component to an offspring generation. In the cause-and-effect scheme, we can determine the life history of an organism using such mechanisms, and whether we ascribe the final cause of a biological individual to perpetuate selfish replicators, or to pursue God as a result of having conscious minds, there is still a question which remains of what moved that first cause. The ultra-reductionist analysis which has been applied to biological individuals has, in effect, asserted that life has some vital force in an attempt to better understand it. However, that there is a greater abundance of the data regarding the physical components making up an organism (and always growing) does not lead us to a holistic understanding of the phenomena that leads to novelty within organisms; that principle which gives life autonomy-without-purpose and causes the structures underlying organisms to transmute into novelty, giving rise to new lineages.
Contrast this account of teleology with the Principia Cybernetica*. The linguistic states that we formulate to study the natural purposes of the living recursively interact with and continually transform/perturb the system under consideration (18). Maturana and Varela say that once an observer actually applies the statement S to their system, the observation made under S will transform subsequent claims made about that system in the future. It is upon this definition that first-order cybernetics bases itself upon. From the point of view of humans, we can see that since Darwin came up with his theory, the natural philosophers that came to follow him operated upon the organisms they studied with the domain of knowledge that Darwin himself interacted in. The idea itself subsequently became absorbed in that recursive process to go on and change what we understood about teleology in the context of life and continues to iterate as modern scientists attempt to apply the theory to lower and lower levels of resolution. As Dennett said, Darwinian evolution has become a universal acid (24), and it is a matter of course that this recursive path it has set us upon will continue to assign meaning to the structures which we study. What S suggests as the generic feature of this teleology is insufficient under the cybernetic view because what we observe as humans will fail to refer to the set of components which are not denoted by structure X but may actually be for a function Y. Although this may sound like solipsism on the surface, I direct the reader towards the instrumentation problem. The structure called “gene” is for the encoding of functional proteins in sexually reproducing life-forms, but in the 19th century the equipment was not available to actually map this structure and establish the gene as a sufficient cause for that clockwork. This failure in reference occurs today with our linear pathways paradigm, particularly in Medicine where advances in Systems Biology are showing associations connections between components that were thought to be completely unrelated before, but are somehow implicated in disease (23). An example of this is the association between the kidney and brain.
However, despite how operationally useful this view of “applying S” may be, there is a paradoxical situation (3) faced by the notion of intrinsic natural purposes. We may only apprehend the natural purposes in nature in terms of a “reciprocal domination” between the parts and the whole of an organism. The poverty of statements such as S lies in the detail that once a final cause has been identified for the parts (and sum-of-parts), then we have in essence already recognized the efficient cause to be inseparable from the final. In essence, the developmental program that we recognize as the intrinsic teleology to interacting agents is precisely that which we want to prove. Van de Vijver states it beautifully by stating that we are “inverting cause and effect in such a way that future events could determine that which happened in an anterior time” (pp. 147). Beginning with that which you set out to prove or otherwise purport is “the end” for a living system is akin to supposing the conclusion of an experiment as its very hypothesis.
Now that it has been shown that organisms have neither immanence nor intrinsic natural purpose based on the interplay between their parts and wholes, I will instead focus on what is meant by teleonomy. I stress first that living systems are without purpose entirely. From a first-order cybernetics point of view, it is my purpose to view such systems from the outside as an observer and integrate teleonomy as a descriptive term for the domain of observations made in the empirical study of the relations underlying the components we can currently handle under the constraints of our technology. Under this framework, there is an end for the living system, and it occurs at the level of the biological individual maintaining itself through its organization (1; pp. 67), although this is unique from the intrinsic purpose which underlies animistic accounts of mechanicism. I consider such an individual to be that which has task closure, which will be expanded on in a later segment. The distinction of observing from the outside versus developing a system within the bounds of say, second-order cybernetics, is the difference of whether we include the observer-community as part of the closure. As was mentioned, it is the case with humans that as we continue observations within a domain of knowledge, we will continue down that research program. By assuming this externalist stance, I seek to extend the current scientific theories we have regarding the study of biological evolution to a generalized view of tackling these largely metaphysical questions regarding where novelty has its origins and how a plurality across biological structures finds itself co-evolving with the external world without purpose.
To address these questions, I introduce entropy. The term entropy has usages within thermodynamics and information theory (7, 13, 15, 17, 19). With respect to being a thermodynamic property, it is the physical quantity which, as it increases over time, determines the ability of life to work and subsequently create useable structures for maintaining its existence (17). In the cell, when the entropy outweighs its capacitive abilities to continue splitting, programmed cell death will occur. With respect to a single living organism, composed of many cells, as this quantity increases over time, order will collapse into ever-increasing disorder as the organism dissipates into other forms of useable energy in the Universe. On the other side, the entropy of information theory (Shannonian) represents a mathematical term for measuring uncertainty (7). This term measures the statistical mechanics of discrete outcomes (15) for a system. One may measure the Shannonian entropy of a coin toss and obtain a solution to how likely it will be to make predictions about its outcome in the future. As the number of coin tosses increases in time, the total entropy begins to increase, implying the unpredictable nature of the outcome as information is lost. Denbigh claims that both of these contexts in which the word is used is independent of man’s presence and thus fully objective on the basis of the persistence of the Universe (pp. 115) and the creativity it manifested pre-human history. Not only is entropy scientifically objective, but the thermodynamic and statistical mechanical forms is also interchangeable within the framework I wish to propose. The basis upon which I consider entropy to be scientifically objective does not necessitate its material existence, however. Rather, it is used here as an abstraction for what appears to be an external phenomena to our observer-community; that as time goes forward, complexity and the irreversibility of the reactions underlying the physico-chemical equations behind life’s self-maintaining processes become greater. It is much easier to reverse the formation of sodium hydroxide than it is to reverse an adult human male back into a fetus. The distinction between reversible and irreversible reactions based on a synthesized view of entropy also allows us to reconsider what is life as it moves from minimal life-forms to incredibly complex ones.
Now that we have a working definition of entropy, I will demonstrate why biological life definitely has a finite initial point. Imagine a machine with a series of knobs on it. This machine (call it B) is controlled by another (A), which works towards optimizing the knobs in the precise positions required for B to begin tinkering with the next machine in the sequence, C. However, if such knobs exist on all the machines in this sequence, then either there is an infinite number of such machines or there was a prime agency which started tinkering with the first machine. In the case of life, if we are to assume the Big Bang occurred 3.5 billion years ago, then organic life cannot be in an infinite state of such machines extending back since time is finite under this theory. In evolutionary time, we know that life on earth did not begin 3.5 billion years ago, as it took an even longer time for the formation of planets even capable of sustaining life. This issue does not even begin to account for the other forms of life which have unfolded asexually or non-genetically. As Brillouin points out, “life is playing upon all these exceptional conditions on the fringe of the second principle of thermodynamics” (pp. 566). Any “prime mover” we know of is residual of how life emerged, although addressing the origin of life cannot give us the “why”. I do not wish to understand the conditions before the seed was planted, only how it lead to the world’s state of affairs as they are today.
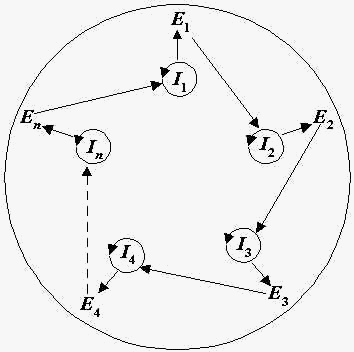
On first-order cybernetics, there have been attempts in the field of theoretical biology to provide an account of organization in living systems using autopoiesis (self-creation) and metabolic-repair (M, R) systems (11). The emergence of rudimentary life-forms has been characterized by the following formula F: (in)organic chemistry => prebiotic metabolism => RNA world => proteins => DNA (13; pp. 1735). The transition between (in)organic chemistry to prebiotic metabolism is particularly important, because the presence of a metabolic cycle is early evidence of rudimentary, self-replicating structures (19) capable of converting useable energy into work (and subsequently, entropy). The earliest work cycles could only have been made possible by what is called task closure. Closure is defined mathematically as a function, or a reference to a function, which has a reference table to all the free variables within the function. To better explain this, consider the term self-referentiality. The clearest example of this is Ouroburos; the world-snake which is forever consuming itself. Self-referentiality in reality is not so extreme, but considers the case of one object, or node, which contains such a pointer back to itself (i.e. the pointer for Ouroburos is his mouth referring back to his tail). When two nodes in concert point to each other, the set over both becomes closed. Experimental and computational evidence has made it quite clear that the existences of such sets are certainly plausible. In the case of that transition between in(organic) chemistry to prebiotic metabolism, metabolic closure based on the Krebs cycle has been proposed as the network underlying minimal life (11). Such reactions are notably reversible, and chemoautotrophs which are living today are capable of this metabolism, which would imply that this lineage has existed from its minimal predecessors. Ashkenazy et al discovered a nine-peptide autocatalytic set capable of achieving closure over a task space which involves the catalysis of all reactions within that set by its very own members out of those nine peptides (21). Others have shown that components such as stem cells can commit to specific lineages based on a rule-based discrete transition at the level of transcriptome-wide noise (2). At what point do these phenomena converge?
Apply the metaphor of machines optimizing one another in sequence to an origin of life scenario on early earth. The scientific literature lends ear to notions such as a primordial soup (a mixture of organic and inorganic components within the open system of the first oceans. Suppose that these first oceans are a finitely bounded, but open, thermal bath (i.e. early earth was “very hot”). Given enough [evolutionary] time (Bywater says it is ~0.3 Gya; pp. 167) the probability of at least one of the organic or inorganic components to collide with one another becomes very high. It is from this collision, similar to the particle collision which unfolded the Universe from the Big Bang that the first primitive relations began, as well as the unfolding of prebiotic, informational components which could self-replicate the very primitive relations implied by that initial contact. Whichever component was first to manifest itself as a “classical” biological structure – the cell membrane, DNA, RNA, cytoplasm – is inconsequential. This is what is meant by component-independence. Instead, if we focus on the primitive relation that emerged from the task closure from minimal life as the efficient cause for biological life to begin on earth, the cybernetic-teleonomic account of life becomes a matter of the change through time of that relation, as implied by the micro-states which it takes, as opposed to a final meaning that is intrinsic to morphological structures. It is on this basis that I pretend to claim Shannonian micro-states perpetuate spontaneous creativity through the production of negative entropy. If the primitive relations underlying the first forms of autonomy and rudimentary life are irreversible, then the living system is guaranteed to also persist and replicate on the basis of entropy in the thermodynamic sense as well (a necessary principle of the second law).
To visualize, suppose that two particles collide in a thermal bath, representing the initial conditions of early earth. Under the collision, the particles form a complex X, which is denoted by the symbol *. The symbol represents the first node (the root) of what is called a binary tree, where successive self-replications of * gives rise to two more, and those two nodes replicate two more nodes and so on. In a sequence, the number of nodes increases in a geometric series on each level, which begins as {1, 2, 4, 8, 16…}. Level n = 0 is defined as the root node. It is at this level that the total information entropy is 1, which represents a finite, yet undetermined, amount of free energy available to convert into useful work within that closure (19). I call this variable ΔI which is the change in information entropy from level n to n + k, where k > 0. In this manner, as the binary tree grows to an indefinite level, the production of negative entropy increases, and the living system loses meaningful micro-states as the information required to maintain the unity begins to experience a loss of integrity. It is at this point that the entropy underlying the relation between the components can no longer undergo self-maintenance and self- renewal, resulting in biological death. When a unique micro-state denoted by ΔI comes about by the behaviour of cells on a continuous space-time plane in two-dimensions, I hypothesize that it is sufficient to be able to trace the emergence of conditions such as cancer. What is especially elegant about tracing such changes through a binary tree is that its growth can be denoted by a logarithmic function, where the logarithm of n (the number of nodes) is actually ΔI As ΔI goes closer to 0, one would expect to observe features such as a loss in genomic stability and a greater number of say, copy-number variations. In the case of n = 0, ΔI = log2 (0) which is 1; intuitively, at the point of origin for an organism, the amount of meaningful information available to its cells to proliferate is 100%. As its cells continue to divide, ΔI becomes smaller and the predictability of what micro-state the cell will take on will continue to grow smaller as well, a result of Shannonian information theory. Within this descriptive domain, the unpredictability of how biological structures, such as the cell membrane, will behave becomes less clear than the statement S which I have originally explained. Evidence from cell signaling studies have shown us that when stem cells move away from their “natural purposes”, the communication between individual cells loses its integrity and as cells within the same tissue are unable to pass messages on to one another, they split into distinct cell populations. As far as a biological program goes, if such a structure as the entire group of cells is for the function of “normal proliferation” leading to non-cancerous tissue, it will inevitably fail as the integrity of the relations between them begins to collapse. At the level of resolution of evolution acting on entire species and sets of individuals constituting a lineage, we can determine their paths insofar as the most probabilistic directions they can take under the constraints imposing themselves on the very micro-states that define those relations, which itself is a recursive process. This does not mean that “the best” structures are selected for based on the solution of an optimization problem, given by the computer gestalt model of evolution (12).
Such autocatalytic sets capable of achieving task closure and performing work (13, 14) are Kantian wholes, where the whole exists for and by the mean of the parts, and the parts for and by the means of the whole (22; pp. 1381). In the context of evolution, the likeliest Kantian wholes to continue to survive in biological evolution were those with the highest probability distribution of obtaining a micro-state x at time t, or the least set of entropy capable of self-maintaining the living functions by means of available, meaningful information within the system. The meaning, behaviours, and even the perceived goal-directedness towards a final cause for organisms are derived from the ways the Kantian wholes assert their autonomy. As rudimentary autocatalytic sets begin to tend towards multicellularity and increase in the total available degrees of freedom (16), the requirement of the living system to develop control systems also begin to manifest themselves. Examples of these are microRNA noise modulators which attenuate DNA coding strands to increase expression levels and the transcription/translation process which is akin to an assembly line for producing functional units (protein). By implementing first-order cybernetics with entropy, I am essentially attributing the intrinsic purposelessness of biological components to their self-referentiality. At the resolution of the Kantian whole, entropy underlies that self-referential pointer of the task closure, and from this foundation of entropy change as an inherently meaningless phenomena (does energy have goals?), I conclude that the only meaning we can assign to the living system is that which emerges from the constraints which a micro-state imposes between components.
However, there are some shortcomings with respect to this view. The first lies with the term entropy itself. I have used it here as a mathematical description – an abstraction – of what seems to happen during the life history of individuals and to account for the origins of novelty. However, even von Neumann, when discussing the matter with Shannon, suggested the term entropy over information or uncertainty for the mere fact that in its thermodynamics context, no one actually knows (and still do not) what entropy actually is as a physical entity in this Universe (7; pp. 113). If I claim that my intention is to nevertheless apply the formalism to biological evolution, and then try to resolve the dynamics of individuals under them, then I have already failed to account for the apparent process of inscribing meanings as an observer embedded within that domain of discourse. Even within the bounds of a second-order cybernetics, where I consider myself as an observer as an input within the feedback loops that underlie living systems, we still cannot say with full confidence that such laws as applied to the thermodynamics and linguistics used to describe those systems – and the subsequent perturbations made by recursive process between the observer (myself) and the individual – apply. Largely this is due to the unresolved gap between the quantum and classical worlds at which evolution enacts upon the biosphere. Furthermore, considering entropy as a primitive relation has been considered by some scientists to be a vital force or perhaps even the res extensa that Descartes describes. However, as I have also recognized, applying this principle of self-referentiality on the level of Kantian wholes does not imply a meaningful substance is where biological individuals derive purposes and intentions from. In the continuous arrow of time, notions of agency, intentionality and meaning – particularly in regards to humans – could be indicative of the discrete transitions under which local compartments (15) emerge given the unique state of entropy flow within that whole. For an example of such a transition, consider the decoupling of the neural tube from the fetus, which gives rise to the brain (6). The development of the brain as such and it becoming an autonomous entity may give rise to inscribed meanings which also lock onto notions of intentions and immanence, but could also be illusions based on the recursions which arise from that “mind” being embedded in the so-called external world. I have derived this line of reasoning as an operational matter constrained within the bounds of the cybernetic-teleonomic account. This cannot be stressed enough, as the self-creating and autonomous nature of the individual exists independently of such first-order cybernetic accounts with which we operate upon them with. To know the inherent purposelessness of life is to understand the mind of God.
REFERENCES
1. Varela, F. (1979) “Principles of Biological Autonomy.” New York: Elsevier North Holland, Inc.
2. Pina, C., Fugazza, C., Tipping, A.J., Brown, J., Soneji, S., Teles, J., Peterson, C., Enver, T.
(2012) “Inferring rules of lineage commitment in hematopoiesis.” Nat Cell Bio, 14(3): 287-94.
3. Van de Vijver, G. (2006) “Kant and the Intuitions of Self-Organization.” Self-Organization and
Emergence in Life Sciences, 143-161.
4. Weber, A., Varela, F. (2002) “Life after Kant: Natural purposes and the autopoietic foundations
of biological individuality.” Phenomenology and the Cognitive Sciences, 1: 97-125.
5. Cummins, R. “Neo-Teleology.” Philosophy of Biology (Part V: 12) in Rosenberg and Arp.
6. Ruiz-Mirazo, K., Moreno, A. (2011) “Autonomy in evolution: from minimal to complex life.”
Synthese, 185: 21-52.
7. Denbigh, K. (1981) “How subjective is entropy?” in Maxwell’s Demon: Entropy Information
Computing, Princeton Series in Physics.
8. Brillouin, L. (1949) “Life, Thermodynamics, and Cybernetics.” in Maxwell’s Demon: Entropy
Information Computing, Princeton Series in Physics.
9. Plato. “Phaedo 99.” Retrieved from Archive.org.
10. Aristotle. “The Organon and Other Works: Physics.” Retrived from Archive.org.
11. Letelier, JC., Marin, G., Mpodozis, J. (2003) “Autopoietic and (M, R) Systems.” J Theor Biol,
2(21): 261-272.
12. Dawkins, R. (1986) “The Blind Watchmaker: The Weasel Program.” Oxford University Press.
13. Hordijk, W., Hein, J., Steel, M. (2010) “Autocatalytic Sets and the Origin of Life.” Entropy, 12:
1733-1742.
14. Monaco, RR., de Montozon, FR. (2012) “Self-Organization, Autocatalysis and Models of the
Origin of Life.” Working draft: NASA Ames Research Center and University for College London.
SUBMITTED.
15. Bennett, CH. (1986) “On the Nature and Origin of Complexity in Discrete, Homogenous,
Locally-Interacting Systems.” Foundations of Physics, 16(6): 585-592.
16. Furusawa, C., Kaneko, K. (2000) “Origin of complexity in multicellular organisms.” Phys Rev
Lett, 84: 6130-6133.
17. Lloyd, S. (1988) “Black Holes, Demons and the Loss of Coherence: How complex systems get
information, and what they do with it.” The Rockefeller University PhD Thesis.
18. Maturana, HR., Varela, F. (1980) “Autopoiesis and Cognition: The Realization of the Living.” D.
Riedel Publishing Company.
19. England, JL. (2012) “Statistical Physics of Self-Replication.” Working draft: MIT Physics
Laboratory. SUBMITTED.
20. Bywater, RP. (2012) “On dating stages in prebiotic chemical evolution.” Naturwissenschaften, 3:
166-176.
21. Zadon, Z, Samiappan, M., Wagner, N., Ashkenasy, N., Ashkenasy, G. (2012) “Peptide Based
Computation: Switches, Gates, and Simple Arithmetic.” Biomol Info Proc: From Logic Sys to
Sensors and Actuators, pp. 9-32.
22. Longo, G., Montevil, M., Kauffman, SA. (2012) “No entailing laws, but enablement in the
evolution of the biosphere.”, Proceedings of the fourth international conference on Genetic and
Evolutionary Computation Conference Companion (GECCO Companion ’12), Terence Soule
(Ed.). ACM, New York, NR, USA, 1379-1392.
23. Huang, S. (2011) “Systems biology of stem cells: three useful perspectives to help overcome the
paradigm of linear pathways.” Philos Trans R Soc Lond B Biol Sci, 366(1575): 2247-59.
24. Dennett, D. (1995) “Darwin’s Dangerous Idea: Evolution and the Meanings of Life.” ISBN: 0-
684-82471-X.
*I derive subsequent orders of cybernetics from this definition as a result of the recursion indicated by the
principle.